

GRAB sensors (G protein-coupled receptor Activation-Based sensors) are the current state of the art for imaging extracellular neurotransmitter dynamics in vivo. These genetically encoded fluorescent indicators detect dopamine, acetylcholine, serotonin, norepinephrine, adenosine, endocannabinoids, and other neuromodulators at the cell surface with high specificity, nanomolar-to-submicromolar affinity, and subsecond temporal resolution — enabling real-time readout of neurotransmitter release in living neurons and intact animal brains.
The Problem GRAB Sensors Solve: Limitations of Classical Neurotransmitter Detection
Understanding how neuromodulators like dopamine and acetylcholine govern behavior requires measuring them where and when they are released — a deceptively difficult problem. Classical approaches each impose a significant compromise:
- Intracerebral microdialysis is the historical gold standard for quantitative extracellular measurements, but its sampling rate is fundamentally slow (typically 1–10 minutes per sample), making it blind to the subsecond dynamics that govern reward signaling and learning. The probe itself is large (70–300 µm in diameter), causing substantial tissue damage and precluding spatially precise measurements (Chefer et al., 2009, Curr Protoc Neurosci; Jaquins-Gerstl and Michael, 2015, Analyst).
- Fast-scan cyclic voltammetry (FSCV) achieves impressive temporal resolution (~10 ms) and sensitivity (~1 nM), but requires electrochemical oxidation for signal generation. This restricts it almost entirely to electroactive catecholamines — primarily dopamine and norepinephrine — and cannot detect acetylcholine, serotonin, or peptide neuromodulators at physiological concentrations. It is also inherently a single-point, single-analyte measurement (Robinson et al., 2008, Chem Rev).
- GCaMP and calcium indicators detect intracellular Ca²⁺ transients as a proxy for neuronal activity and neurotransmitter release, but calcium is a downstream signal — it does not report which neurotransmitter was released, nor its concentration in the extracellular space or synaptic cleft. Calcium imaging cannot distinguish between glutamate release and dopamine release from the same cell.
GRAB sensors fill this gap. Because they are genetically encoded, they can be delivered to specific cell types via AAV vectors, expressed on the neuronal plasma membrane, and imaged using the same fluorescence hardware already in use for GCaMP experiments. They detect specific neurotransmitters in the extracellular space — not as a proxy, but directly — with millisecond-to-second temporal resolution and single-cell or better spatial precision.
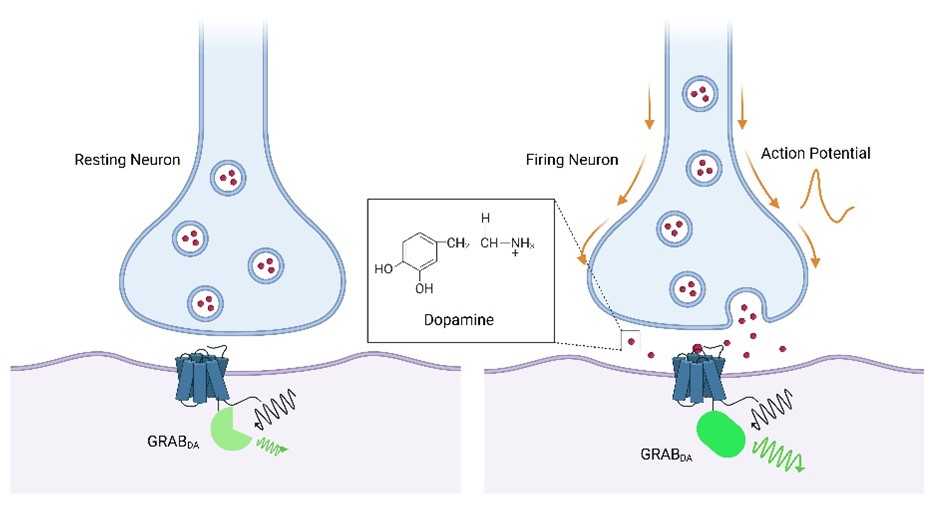
GRAB Sensor Mechanism: How They Work
The GRAB sensor platform was originally developed by the Yulong Li laboratory at Peking University and is the defining contribution to genetically encoded neuromodulator sensing. The core engineering principle is elegant and generalizable across dozens of neurotransmitter systems.
Architecture. A circularly permuted GFP (cpGFP) — the same conformation-sensitive fluorescent protein used in GCaMP calcium indicators — is inserted into the third intracellular loop (ICL3) of a selected human G protein-coupled receptor (GPCR). The GPCR provides the ligand-binding specificity; the cpGFP reports the conformational change.
Signal generation. When the target neurotransmitter binds to the extracellular domain of the GPCR, the receptor undergoes its characteristic conformational rearrangement. This structural change is transduced through the receptor backbone to the ICL3-embedded cpGFP, altering the chromophore environment and producing a robust increase in fluorescence intensity (ΔF/F). The sensor thus converts binding of an extracellular molecule into an optical signal detectable by standard fluorescence microscopy.
Critically, GRAB sensors do not activate downstream G protein signaling. The ICL3 insertion disrupts the G protein coupling interface, and pharmacological validation has confirmed that expression of GRAB sensors — for example, GRAB_DA in response to dopamine — does not produce changes in intracellular cAMP, does not trigger β-arrestin recruitment, and does not cause receptor internalization. The sensor acts as a pure reporter: it reads neurotransmitter concentration at the cell surface without perturbing the neuron's physiology (Sun et al., 2020, Nat Methods).
Distinction from iGluSnFR. It is important to note that iGluSnFR (intensity-based glutamate-sensing fluorescent reporter) is not a GRAB sensor. iGluSnFR uses a completely different scaffold: a bacterial periplasmic binding protein from E. coli (GltI) fused to cpGFP (Marvin et al., 2013, Nat Methods). It detects extracellular glutamate with excellent performance and is often used alongside GRAB sensors, but the two platforms are architecturally distinct. GRAB sensors are GPCR-based; iGluSnFR and iGABASnFR are bacterial periplasmic binding protein-based.
Available GRAB Sensors: Comprehensive Reference Table
The following table lists published GRAB sensors and related genetically encoded neuromodulator indicators in current use. Sensor naming follows the convention that number suffixes denote generation (higher = improved performance) and letter suffixes denote affinity: h = high affinity, m = moderate affinity, l = low affinity. A mut (mutant) variant is a ligand-insensitive control used to verify signal specificity. Newer generations typically offer improved dynamic range, faster kinetics, or better photostability compared to earlier versions.
| Sensor | Target | Fluorophore | Peak Ex / Em | Key Reference |
|---|---|---|---|---|
| GRAB_ACh3.0 (GACh3.0) | Acetylcholine | cpEGFP (green) | ~490 nm / ~510 nm | Jing et al., 2018, Nat Biotechnol |
| GRAB_DA2m / GRAB_DA2h | Dopamine | cpEGFP (green) | ~490 nm / ~510 nm | Sun et al., 2020, Nat Methods |
| GRAB_DA3m / GRAB_DA3h | Dopamine (3rd generation) | cpEGFP (green) | ~490 nm / ~510 nm | Sun et al., 2020, Nat Methods |
| rGRAB_DA1m / rGRAB_DA1h (rDA) | Dopamine (red) | cpmApple (red) | ~560 nm / ~600 nm | Sun et al., 2020, Nat Methods |
| GRAB_NE1m (GRAB_NE) | Norepinephrine | cpEGFP (green) | ~490 nm / ~510 nm | Feng et al., 2019, Neuron |
| GRAB_5-HT1.0 / GRAB_5-HT3.0 | Serotonin (5-HT) | cpEGFP (green) | ~490 nm / ~510 nm | Wan et al., 2021, Nat Neurosci |
| GRAB_eCB2.0 | Endocannabinoids (2-AG, AEA) | cpEGFP (green) | ~490 nm / ~510 nm | Dong et al., 2022, Nat Biotechnol |
| iGluSnFR3 (non-GRAB) | Glutamate (extracellular) | cpEGFP (green) | ~490 nm / ~510 nm | Marvin et al., 2013, Nat Methods |
Note: iGluSnFR and iGABASnFR are bacterial periplasmic binding protein-based sensors, not GRAB sensors. They are listed here for reference because they are commonly used alongside GRAB sensors in multicolor experiments. Check the BioHippo AAV catalog for the most current sensor generations available as ready-to-use virus.
Experimental Setup: Using GRAB Sensors in Practice
GRAB sensors are delivered and expressed using the same AAV workflow as GCaMP and other genetically encoded indicators. The sensor plasmid is packaged into an AAV capsid and injected stereotaxically into the brain region of interest.
AAV Delivery
- Serotype: AAV9 and AAV-PHP.eB are the most commonly used serotypes for neuronal GRAB sensor expression. AAV9 provides robust pan-neuronal transduction after stereotaxic injection; PHP.eB achieves broader transduction after systemic delivery in mice. rAAV2-retro can be used for retrograde labeling of projection neurons. Available serotypes via BioHippo AAV catalog.
- Promoter: hSyn (human synapsin-1) drives pan-neuronal expression and is the standard choice for most GRAB sensor experiments. EF1α or CAG are used when broader or stronger expression is needed. Cre-dependent constructs (DIO/FLEX) allow cell-type-specific targeting when combined with a Cre-driver mouse line.
- Expression window: Allow 2–4 weeks post-injection for peak membrane expression of the sensor before beginning imaging experiments. Long-term expression is stable: robust signals have been reported 3–6 months after AAV injection in mice.
- Titer: Use titers of ≥2 × 10¹² vg/mL for reliable in vivo expression. BioHippo GRAB sensor AAVs are supplied at ≥2 × 10¹² vg/mL.
Imaging Modalities
- Fiber photometry is the most widely used readout for GRAB sensors in freely behaving animals. A fiber optic cannula implanted above or into the target region collects population-level ΔF/F at 10–100 Hz sampling rates. It provides excellent sensitivity for detecting neurotransmitter dynamics during behavior (locomotion, reward, fear conditioning, social interaction) but does not resolve individual cells or axonal compartments.
- Two-photon (2P) microscopy provides single-cell and sub-cellular spatial resolution, resolving individual dendritic spines, axonal boutons, and single synaptic release events. It requires a cranial window preparation and a head-fixed preparation, making it best suited to superficial cortical regions. Two-photon excitation minimizes phototoxicity and background fluorescence in thick tissue.
- Widefield epifluorescence (mesoscale imaging) allows whole-dorsal-cortex imaging at 10–30 Hz frame rates using a macro-objective. It trades resolution for field of view and is well suited to mapping neurotransmitter dynamics across large cortical areas simultaneously.
- Miniscope (1P head-mounted microscope) enables single-cell imaging of GRAB sensor fluorescence in deep brain structures (striatum, hippocampus, amygdala) in freely moving animals. Resolution is lower than two-photon but compatible with complex behavioral paradigms.
Essential Controls
- Ligand-insensitive mutant control: Every major GRAB sensor family includes a validated pharmacologically insensitive mutant (e.g., GRAB_DA_mut, GACh_mut). Co-inject the mutant in a separate animal or brain region and confirm that it produces no fluorescence response to the cognate neurotransmitter. This is the most important specificity control.
- Pharmacological validation: Apply specific agonists and antagonists (e.g., D1R/D2R agonists and antagonists for GRAB_DA; atropine for GRAB_ACh) to confirm that the fluorescence signal is blocked by the cognate receptor antagonist and mimicked by the agonist.
- Dual-color imaging: Co-express a spectrally separated calcium indicator (jRGECO1a, RCaMP2, or jRCaMP1a — red channel) alongside the green GRAB sensor to simultaneously record neurotransmitter release and upstream or downstream neuronal calcium activity in the same experiment.
Combining GRAB Sensors with Optogenetics and DREADDs
GRAB sensors are fully compatible with all-optical interrogation strategies. Because the most widely used GRAB sensors use green fluorescent protein (cpEGFP, Ex ~490 nm), they can be paired with red-shifted channelrhodopsins and opsins (ChrimsonR, ChRmine, Chrimson) that are activated by wavelengths (>590 nm) that do not excite cpEGFP. This spectral separation enables true all-optical experiments: stimulate a dopamine-releasing circuit with red light, then simultaneously read out dopamine release at the target projection using green GRAB_DA fluorescence.
For DREADD experiments, chemogenetic stimulation of a defined neuronal population (e.g., CNO activation of hM3Dq in dopamine neurons) followed by GRAB sensor readout in the projection target provides a clean, non-invasive way to confirm that the chemogenetic manipulation actually drove neurotransmitter release. This approach has been used to characterize dopaminergic, noradrenergic, and cholinergic circuit function in intact behaving animals.
Red fluorescent GRAB sensors (rGRAB_DA, rGRAB_ACh) expand the palette further: they can be imaged in the red channel while green calcium indicators (GCaMP variants) or green channelrhodopsins (ChR2) are used in the green channel, enabling orthogonal readout of neurotransmitter dynamics and neuronal activity from the same preparation without spectral crosstalk.
Ready-to-Use GRAB Sensor AAVs at BioHippo
BioHippo stocks a comprehensive library of ready-to-use GRAB sensor AAVs, including the latest generations of dopamine (GRAB_DA3m, GRAB_DA3h, rGRAB_DA), acetylcholine (GACh/GRAB_ACh), norepinephrine (GRAB_NE), serotonin (GRAB_5-HT), and endocannabinoid (GRAB_eCB) sensors. All constructs are available across a wide range of serotypes (AAV2/1, AAV2/2, AAV2/5, AAV2/8, AAV2/9, PHP.eB, rAAV2-retro, and others) under hSyn, CAG, EF1α, and CMV promoters. Constitutive and Cre-dependent (DIO/FLEX) versions are available to support cell-type-specific targeting.
All BioHippo AAVs are supplied at ≥2 × 10¹² vg/mL in 100 µL aliquots. Custom packaging — including non-listed sensor versions, alternate promoters, or novel serotypes — is available on request. To browse the full GRAB sensor AAV catalog or request a quote, contact orders@biohippo.com.
New to AAV? See our primer: New to AAV? Here Are the Things You Need to Know.
Frequently Asked Questions
What is a GRAB sensor?
A GRAB sensor (G protein-coupled receptor Activation-Based sensor) is a genetically encoded fluorescent indicator that detects specific extracellular neurotransmitters in real time. It consists of a circularly permuted GFP (cpGFP) inserted into the intracellular loop of a human GPCR. When the target neurotransmitter binds to the receptor's extracellular domain, the resulting conformational change increases cpGFP fluorescence, providing a direct optical readout of neurotransmitter concentration at the cell surface. GRAB sensors were developed by the Yulong Li laboratory at Peking University and cover dopamine, acetylcholine, norepinephrine, serotonin, adenosine, endocannabinoids, and other neuromodulators.
How are GRAB sensors different from GCaMP calcium indicators?
GCaMP indicators detect intracellular Ca²⁺ transients as a proxy for neuronal firing and neurotransmitter release from the expressing cell. GRAB sensors detect specific neurotransmitters in the extracellular space — at the cell surface of the sensor-expressing neuron. This means GRAB sensors can identify which neurotransmitter is present and estimate its local concentration, which GCaMP cannot. A neuron expressing GCaMP reports its own activity; a neuron expressing GRAB_DA reports how much dopamine is arriving from dopaminergic afferents. The two sensor types are fully compatible: they can be co-expressed in different channels for simultaneous readout of input neurotransmitter release and the host cell's calcium response.
What AAV do I use to express GRAB sensors in neurons?
For most in vivo neuroscience experiments, AAV9 or AAV-PHP.eB under the hSyn (human synapsin-1) promoter is the standard choice for pan-neuronal GRAB sensor expression. PHP.eB achieves broader CNS transduction after systemic (IV) delivery in mice. For deep brain targets in rats or non-human primates, AAV9 via stereotaxic injection is recommended. Cre-dependent (DIO/FLEX) constructs in combination with a cell-type-specific Cre-driver line restrict expression to defined cell populations (e.g., D1-Cre or D2-Cre mice for dopamine receptor subtype-specific expression). Allow 2–4 weeks post-injection for peak expression before beginning imaging.
Can I combine GRAB sensors with optogenetics?
Yes. GRAB sensors (typically green, cpEGFP-based) are spectrally compatible with red-shifted channelrhodopsins such as ChrimsonR and ChRmine, which are excited by red light (>590 nm) that does not activate cpEGFP. This enables all-optical experiments: stimulate a projection with red-shifted opsin while simultaneously imaging neurotransmitter release with green GRAB sensor. For experiments requiring green-channel optogenetics (ChR2), use red fluorescent GRAB variants (rGRAB_DA, rGRAB_ACh) that can be imaged in the red channel without spectral interference.
Are GRAB sensors available as ready-to-use AAV?
Yes. BioHippo stocks a comprehensive library of GRAB sensor AAVs — including GRAB_DA, GRAB_ACh, GRAB_NE, GRAB_5-HT, GRAB_eCB, and their red-fluorescent and mutant control variants — across more than 20 serotypes. All are supplied at ≥2 × 10¹² vg/mL in 100 µL aliquots for immediate use. Browse the full list in the BioHippo AAV collection or contact orders@biohippo.com for custom packaging requests.
What do the abbreviations h, m, l, and mut mean in GRAB sensor names?
These letter suffixes denote the sensor's apparent affinity for its target ligand: h = high affinity (lower K_D, better for detecting low neurotransmitter concentrations), m = moderate affinity (suited to intermediate concentration ranges), l = low affinity (suited to high concentration ranges or avoiding saturation during strong stimulation). mut indicates a ligand-insensitive mutant version used as a pharmacological specificity control — it shares all structural features of the active sensor but does not respond to the cognate neurotransmitter. Number suffixes (e.g., DA2m vs. DA3m) denote generation; higher numbers indicate improved performance (greater dynamic range, faster kinetics, or improved photostability).
Can GRAB sensors be expressed in vivo long term?
Yes. AAV-mediated GRAB sensor expression in mouse brain is stable long term. Reliable fluorescence signals have been recorded from GRAB sensor-expressing neurons 3–6 months after AAV injection, with no reported signs of cytotoxicity, altered membrane potential, or changes in synaptic responses in validated preparations (Jing et al., 2018; Feng et al., 2019).
References
Chefer VI, Thompson AC, Zapata A, Shippenberg TS. Overview of brain microdialysis. Curr Protoc Neurosci. 2009;Chapter 7:Unit7.1. doi: 10.1002/cpns.5. PMID: 19340812.
Dong A, He K, Dudok B, Farrell JS, et al. A fluorescent sensor for spatiotemporally resolved imaging of endocannabinoid dynamics in vivo. Nat Biotechnol. 2022;40(5):787–798. doi: 10.1038/s41587-021-01074-4. PMID: 34764491.
Feng J, Zhang C, Lischinsky JE, Jing M, et al. A genetically encoded fluorescent sensor for rapid and specific in vivo detection of norepinephrine. Neuron. 2019;102(4):745–761.e8. doi: 10.1016/j.neuron.2019.02.037. PMID: 30922875.
Jaquins-Gerstl A, Michael AC. A review of the effects of FSCV and microdialysis measurements on dopamine release in the surrounding tissue. Analyst. 2015;140(11):3696–3708. doi: 10.1039/c5an00087d.
Jing M, Zhang P, Wang G, Feng J, et al. A genetically encoded fluorescent acetylcholine indicator for in vitro and in vivo studies. Nat Biotechnol. 2018;36(8):726–737. doi: 10.1038/nbt.4184. PMID: 29985477.
Marvin JS, Borghuis BG, Tian L, Cichon J, et al. An optimized fluorescent probe for visualizing glutamate neurotransmission. Nat Methods. 2013;10(2):162–170. doi: 10.1038/nmeth.2333. PMID: 23314171.
Robinson DL, Hermans A, Seipel AT, Wightman RM. Monitoring rapid chemical communication in the brain. Chem Rev. 2008;108(7):2554–2584. doi: 10.1021/cr068081q. PMID: 18576692.
Sun F, Zhou J, Dai B, Qian T, et al. Next-generation GRAB sensors for monitoring dopaminergic activity in vivo. Nat Methods. 2020;17(11):1156–1166. doi: 10.1038/s41592-020-00981-9. PMID: 33087905.
Wan J, Peng W, Li X, Qian T, et al. A genetically encoded sensor for measuring serotonin dynamics. Nat Neurosci. 2021;24(5):746–752. doi: 10.1038/s41593-021-00823-7. PMID: 33821000.